The process by which cells leave the bloodstream and cross the endothelium to enter into various tissues is called extravasation. Although the particular molecules involved may differ in different situations, the fundamental process is the same. Extravasation can be divided into three stages – rolling, activation and firm attachment, and trans-endothelial migration. Once cells have left the bloodstream they must be guided to the right location within the tissue. The entrance of neutrophils into a site of inflammation is the best understood example and will be described to illustrate the basic steps involved in these processes.
1. Rolling. Neutrophils, like other leukocytes, normally travel in the centre of the blood flow away from the endothelium. At a site of inflammation asodilation occurs, slowing down and disturbing the blood flow so that the neutrophils can ‘bump’ along the endothelium, a process known as rolling. Due to the action of inflammatory mediators, especially TNFα, the endothelial cells are activated to express P-selectin and E-selectin on their surface. These selectins can bind to sialyl-Lewisx on the surface of the neutrophil, slowing down the neutrophil so that it rolls along the endothelium.
2. Activation and firm attachment. The binding of the selectins to the sialyl-Lewisx is not strong enough for the neutrophil to adhere strongly to the endothelium. Strong attachment requires the binding of the integrin LFA-1 on the neutrophil to ICAM-1 on the endothelium. Before it can bind to ICAM-1, the LFA-1 must change conformation. One of the factors produced in an inflammatory response is interleukin- 8 (IL-8), which is a chemokine. Chemokines are a group of cytokines with chemotactic and other functions. Some of the IL-8 produced is held in the extracellular matrix on the endothelial cell surface and can bind to IL-8 receptors on the neutrophil surface. The binding of IL-8 to the neutrophil activates the neutrophil and LFA-1 changes conformation and binds firmly to ICAM-1 on the endothelium.
3. Transendothelial migration. Once the neutrophil is firmly attached to the endothelium it squeezes between the endothelial cells, making contact with the basement membrane underneath. This process is poorly understood but involves additional adhesion molecules. Finally enzymes digest the basement membrane, allowing the leucocyte to pass through into the tissue space.
4. Movement in the site of inflammation. In the inflamed tissue there will be a gradient of IL-8, with maximum levels at the centre of infection. Neutrophils that have left the bloodstream and entered the tissue will travel along the IL-8 gradient, moving towards increasing concentration of the chemokine so that they will accumulate at the centre of infection.
The way in which other leukocytes cross endothelia, leave the bloodstream and migrate through tissues is essentially the same as for neutrophils, although the adhesion molecules and chemokines may be different for different cell types. Many adhesion molecules and chemokines exist to control adhesion, integrin activation and movement of different types of cells in various tissues. In sites of inflammation other factors such as complement components and prostaglandins can also act as chemoattractants.
понедельник, 1 ноября 2010 г.
суббота, 30 октября 2010 г.
Adhesion molecules.
There are four families of adhesion molecules called selectins, integrins, mucin-like vascular addressins and members of the immunoglobulin superfamily and each family contains many members. Different adhesion molecules bind to each other in a specific manner and enable cells to interact with each other. Cell–cell adhesion is controlled both by the expression of particular adhesion molecules and in some cases by the activation status, or actual binding capacity, of the adhesion molecules.

Different adhesion molecules are expressed on different cell types; some are expressed constantly on the cell surface and others are induced by cell activation, e.g. by cytokines.
By altering cell-adhesion molecule expression or activity on endothelial cells or leukocytes, it is possible to control whether particular leukocytes bind to endothelium at a particular tissue site and, hence, the entry of the leukocytes into the tissue.
Selectins are glycoproteins that are lectins, i.e. sugar-binding molecules, some of which are expressed on leukocytes and some on endothelial cells.
Mucin-like vascular addressins are heavily glycosylated proteins and therefore can bind to the selectins. Some are expressed on leukocytes and some on endothelial cells.
Integrins are heterodimeric proteins consisting of an α-chain and a β- chain and are expressed on leukocytes. There are many α- and β-chains and they can pair to give many combinations of integrins with different expression and binding specificity. Some integrins will bind to target molecules only following activation of the leukocyte by various factors.
Immunoglobulin superfamily: these molecules contain immunoglobulin (Ig)-like domains (110 amino acids flanked by an intra-chain disulphide bond) and are the binding target for the integrins. They are expressed on endothelial cells.

Adhesion molecules.
Different adhesion molecules are expressed on different cell types; some are expressed constantly on the cell surface and others are induced by cell activation, e.g. by cytokines.
By altering cell-adhesion molecule expression or activity on endothelial cells or leukocytes, it is possible to control whether particular leukocytes bind to endothelium at a particular tissue site and, hence, the entry of the leukocytes into the tissue.
Selectins are glycoproteins that are lectins, i.e. sugar-binding molecules, some of which are expressed on leukocytes and some on endothelial cells.
Mucin-like vascular addressins are heavily glycosylated proteins and therefore can bind to the selectins. Some are expressed on leukocytes and some on endothelial cells.
Integrins are heterodimeric proteins consisting of an α-chain and a β- chain and are expressed on leukocytes. There are many α- and β-chains and they can pair to give many combinations of integrins with different expression and binding specificity. Some integrins will bind to target molecules only following activation of the leukocyte by various factors.
Immunoglobulin superfamily: these molecules contain immunoglobulin (Ig)-like domains (110 amino acids flanked by an intra-chain disulphide bond) and are the binding target for the integrins. They are expressed on endothelial cells.
пятница, 29 октября 2010 г.
Cell migration.
The movement of cells around the body must be carefully controlled so that the cells go only to where they are required. This control is at two levels: one level is controlling where leukocytes leave the bloodstream; the second level is controlling where the cells go within tissues and organs once they have left the bloodstream. For a single cell, most organs are pretty big places and the cell must go to the right location within the organ or tissue.
Two important factors play an important role in controlling the movement of cells to and within specific tissue sites. Adhesion molecules are present on leucocytes and endothelial cells, and interactions between adhesion molecules allow leucocytes to bind to endothelium as part of the process of migrating across the endothelium. Chemotactic agents, especially the chemokines, are also important in controlling cell migration. They can act directly on cells and cause them to move in a particular direction or they can act indirectly by altering the expression or binding activity of adhesion molecules.
Two important factors play an important role in controlling the movement of cells to and within specific tissue sites. Adhesion molecules are present on leucocytes and endothelial cells, and interactions between adhesion molecules allow leucocytes to bind to endothelium as part of the process of migrating across the endothelium. Chemotactic agents, especially the chemokines, are also important in controlling cell migration. They can act directly on cells and cause them to move in a particular direction or they can act indirectly by altering the expression or binding activity of adhesion molecules.
The inflammatory response and cell migration.
If a pathogen has successfully invaded a tissue, the macrophages in the tissue may recognise the pathogens with one of the receptors and attempt to phagocytose and kill the pathogens. Often there are not enough macrophages present in a tissue to phagocytose and remove all the pathogens and therefore the tissue macrophages initiate a response that will bring additional phagocytes, together with a variety of proteins, to the site of infection from the blood. These cells and proteins then help to remove the pathogen. This response is known as the inflammatory response. The aim of the inflammatory response is to recruit cells and other factors from the bloodstream into tissues to aid in the remove of pathogens and dead cells or tissue. Leukocytes (white blood cells) are unique in their ability to move throughout the body. They travel through the bloodstream and also have the ability to leave the bloodstream and enter tissue or organs. This ability to move around the body is also referred to as ‘cell migration’.
четверг, 28 октября 2010 г.
Cytokines.
The term cytokine covers a large number of smallish proteins (usually less than 20 kDa) that serve a hormone-like function in enabling cells to communicate with each other. Most people are familiar with hormones such as insulin and growth hormone, which are produced in one organ or tissue and travel through the bloodstream to other organs where they bind to receptors on the cells of that organ and stimulate a particular response. Hormones that are produced in one organ and act on a distant tissue are said to be acting in an endocrine manner. Cytokines do not usually act in an endocrine manner; rather, they act locally. They are produced by cells in a particular tissue and act on ‘cells’ in that tissue. Cytokines therefore act in a paracrine or autocrine manner. Paracrine action means that the cytokine binds to receptors on cells close to those producing the cytokine; by ‘close’ we are probably talking about a microenvironment of microns to 1 mm. Autocrine means that the cytokine actually binds to receptors on the cell that produced the cytokine. Thus the role of cytokines is to enable cells to communicate with each other in a local environment. A few cytokines can also act in an endocrine manner.
There are many cytokines and they can be divided into families. The main families of cytokines are the interleukins, colony-stimulating G-CSF, granulocyte-CSF; M-CSF, macrophage-CSF; GM-CSF, granulocyte/monocyte-CSF; MCP, macrophage chemotactic protein; TGF, transforming growth factor; IGF, insulin-like growth factor.

The functions of cytokines will be described in detail at the appropriate times when particular mechanisms are being explained. It is important to realise that in the body, cells are never exposed to a single cytokine – they will be exposed to a number of different cytokines, probably produced by a number of different cell types. Different cytokines can either act cooperatively in promoting a response or act antagonistically in inhibiting each other’s actions. It is the combination of cytokines to which a cell is exposed that determines the behaviour of the cell.
There are many cytokines and they can be divided into families. The main families of cytokines are the interleukins, colony-stimulating G-CSF, granulocyte-CSF; M-CSF, macrophage-CSF; GM-CSF, granulocyte/monocyte-CSF; MCP, macrophage chemotactic protein; TGF, transforming growth factor; IGF, insulin-like growth factor.

Action of hormones. Endocrine: the hormone is secreted at one site of the body and travels through the bloodstream. The hormone will bind to receptors on cells at a distant site (blue cells) and cause a response in those cells. Paracrine: hormones produced by cells in a tissue bind to receptors on other cells in the immediate vicinity (blue cells). Cells in other parts of the same tissue are not affected by the hormone (white cells). Autocrine: the secreted hormone binds to receptors on the cell that produced the hormone (blue cell).
The functions of cytokines will be described in detail at the appropriate times when particular mechanisms are being explained. It is important to realise that in the body, cells are never exposed to a single cytokine – they will be exposed to a number of different cytokines, probably produced by a number of different cell types. Different cytokines can either act cooperatively in promoting a response or act antagonistically in inhibiting each other’s actions. It is the combination of cytokines to which a cell is exposed that determines the behaviour of the cell.
The cellular response to infection – production of new factors.
When cells of the innate immune system encounter pathogenic products they can respond in ways other than phagocytosis. Cells can be stimulated to synthesise and/or secrete an enormous variety of new products. Some of these products may be directly involved in killing pathogens. Other products are involved indirectly in recruiting other cell types to try and eliminate the pathogen. An important group of proteins that can be secreted in response to pathogenic stimuli are known as cytokines.


Recognition by phagocytes. Phagocytes must distinguish microbes and dead host cells from healthy host cells so that healthy host cells are not phagocytosed. Phagocytes have receptors on their surface that recognise sugars present on microbes or sugars that are newly expressed on dead or damaged host cells. These sugars are not present on healthy host cells and therefore the host cells are not phagocytosed.
суббота, 23 октября 2010 г.
The cellular response to infection – phagocytosis.
Phagocytosis is the ingestion and destruction of microbes by cells called phagocytes. The two main types of phagocytes are the macrophages and neutrophils described above. The way in which macrophages and neutrophils phagocytose particles is essentially the same and can be divided into four stages.
1. Attachment of the phagocyte to the particle being phagocytosed, which may be a pathogen, a dead or damaged host cell or a piece of tissue.
2. Ingestion. By extending membrane protrusions called pseudopodia around the particle, the phagocyte is able to engulf the particle, which is taken into the cell in a phagocytic vacuole.
3. Killing. If the ingested particle is a live cell of a pathogen (e.g. a bacterium) the phagocyte will normally kill the cell by one of a number of mechanisms.
4. Degradation. The phagocytosed particle, whether it is a dead cell or a piece of tissue, is broken down by enzymes in the phagocytic vacuole.

Although the basic process of phagocytosis is similar in neutrophils and macrophages, there is an important difference. While neutrophils are only able to phagocytose small organisms such as bacteria and viruses, macrophages are able to phagocytose larger particles such as dead cells and tissue debris in addition to microorganisms. Therefore macrophages are involved in eliminating pathogens from tissues and also in cleaning up damaged tissue by removing dead or damaged host cells. Macrophages are able to distinguish between healthy host cells and dead/damaged cells because the receptors they have for recognising sugars on microbes also recognise sugars that are exposed by dead or damaged host cells.
1. Attachment of the phagocyte to the particle being phagocytosed, which may be a pathogen, a dead or damaged host cell or a piece of tissue.
2. Ingestion. By extending membrane protrusions called pseudopodia around the particle, the phagocyte is able to engulf the particle, which is taken into the cell in a phagocytic vacuole.
3. Killing. If the ingested particle is a live cell of a pathogen (e.g. a bacterium) the phagocyte will normally kill the cell by one of a number of mechanisms.
4. Degradation. The phagocytosed particle, whether it is a dead cell or a piece of tissue, is broken down by enzymes in the phagocytic vacuole.

Phagocytosis. Phagocytes can take up and remove bacteria and dead host cells or tissue debris. The figure shows phagocytosis of a bacterium: ➀ The phagocyte binds to the bacterium. ➁ The phagocyte extends projections around the bacterium and engulfs it in a phagocytic vacuole. ➂ The phagocyte kills the engulfed bacterium. ➃ The bacterium is degraded by proteolytic enzymes.
Although the basic process of phagocytosis is similar in neutrophils and macrophages, there is an important difference. While neutrophils are only able to phagocytose small organisms such as bacteria and viruses, macrophages are able to phagocytose larger particles such as dead cells and tissue debris in addition to microorganisms. Therefore macrophages are involved in eliminating pathogens from tissues and also in cleaning up damaged tissue by removing dead or damaged host cells. Macrophages are able to distinguish between healthy host cells and dead/damaged cells because the receptors they have for recognising sugars on microbes also recognise sugars that are exposed by dead or damaged host cells.
пятница, 22 октября 2010 г.
The cellular response to recognition of microbial products.
It can be seen that many cell types of the innate immune system have receptors on their surface that can recognise microbial products and therefore detect that we have been infected. Obviously recognition of infection is useful only if it results in a response to the infection that will ideally eliminate the pathogenic organism or at the very least limit the replication and spread of the pathogen. There are a number of ways in which different cells can respond to recognition of pathogens, and one of these is phagocytosis.



Phagocyte receptors

Recognition by Toll-like receptors
вторник, 19 октября 2010 г.
Recognition of pathogens by cells of the innate immune system.
When we say that cells of the innate immune system can ‘recognise’ pathogens, what do we actually mean? By recognition we mean that molecules, or receptors, on cells of the innate immune system bind to other molecules that are present on pathogens but not present on our own cells. In this way the immune system can distinguish foreign objects and respond to a foreign pathogen but not respond against our own tissue or cells. It is a crucial feature of the immune system that it can target foreign objects. There are many receptors present on cells of the innate immune system and some.
An important family of receptors that are able to recognise a wide variety of pathogen-associated molecules has been identified. These are the Toll-like receptors (TLRs), which were named because of their structural relationship to Toll, a protein involved in development of Drosophila, the fruit fly. There are about 10 TLRs that have been identified in man and similar numbers in other species. The TLRs recognise a variety of microbial products such as doublestranded RNA (found only in viral infection), lipopolysaccharide (LPS) from bacterial cell walls, bacterial lipoproteins, unmethylated DNA (indicating it is of bacterial origin) and flagellin, a component of bacterial flagella. Different TLRs recognise different microbial products.
An important family of receptors that are able to recognise a wide variety of pathogen-associated molecules has been identified. These are the Toll-like receptors (TLRs), which were named because of their structural relationship to Toll, a protein involved in development of Drosophila, the fruit fly. There are about 10 TLRs that have been identified in man and similar numbers in other species. The TLRs recognise a variety of microbial products such as doublestranded RNA (found only in viral infection), lipopolysaccharide (LPS) from bacterial cell walls, bacterial lipoproteins, unmethylated DNA (indicating it is of bacterial origin) and flagellin, a component of bacterial flagella. Different TLRs recognise different microbial products.
понедельник, 18 октября 2010 г.
Cells of the innate immune system.
There are also many cells and proteins found throughout the body that are part of the innate immune system and are involved in defence against pathogens. Collectively these cells and proteins perform two important functions: they are able to recognise the presence of a foreign entity, for example a bacterial infection, and they provide an immediate cellular response to the presence of an infectious agent. The nature of this cellular response differs according to the nature of the cell and the way in which it is stimulated.
One important cell type of the innate immune system is the macrophage, which is a bone marrow-derived cell type found in most tissues. Some macrophages reside for a long time in the same tissue and are therefore called fixed macrophages. Other macrophages live up to their name of wandering macrophages by moving through tissue. Macrophages are related to a type of white blood cell called a monocyte, which is also derived from bone marrow. Monocytes can leave the bloodstream and enter tissue, where they differentiate into macrophages. Because of their relationship, macrophages and monocytes are known as cells of the monocyte/macrophage lineage.
One important cell type of the innate immune system is the macrophage, which is a bone marrow-derived cell type found in most tissues. Some macrophages reside for a long time in the same tissue and are therefore called fixed macrophages. Other macrophages live up to their name of wandering macrophages by moving through tissue. Macrophages are related to a type of white blood cell called a monocyte, which is also derived from bone marrow. Monocytes can leave the bloodstream and enter tissue, where they differentiate into macrophages. Because of their relationship, macrophages and monocytes are known as cells of the monocyte/macrophage lineage.
пятница, 15 октября 2010 г.
The immediate response to infection – the innate immune system.
The term ‘innate immune system’ is used to describe pre-existing defence mechanisms that are designed to prevent infection by pathogens or to mount an immediate defence against the infectious agent. The physical, chemical and biochemical barriers to infection are part of the innate immune system. They are called ‘innate’ or ‘natural’ because they are present before infection, although the amount of some components may increase following infection.
The response to infection.
Covered pathogenic organisms and the stages of infection leading to disease, illustrating the tremendous variety of organisms and disease processes to which the body is exposed. Now we turn to the problem of how the body attempts to counter the threats posed by this enormous number of different infectious organisms.
The response to infection can be divided into five stages:
1. Awareness of infection. Obviously the body cannot begin to mount a defensive response against a pathogen until it is aware of the presence of the pathogen.
2. The immediate response to infection. This involves the activity of cells and other factors that are present at the time of infection but may require their recruitment to the site of infection and activation once there.
3. The delayed response to infection. If the immediate response to infection is not effective in eliminating the pathogen, the next stage is to generate new cells and factors to deal with the infection.
4. Destruction or elimination of the pathogen or neutralisation of the threat posed by pathogens. The optimal way of dealing with pathogens is to kill them or eliminate them from the body. However, in some cases where pathogens are producing a powerful toxin it may be more beneficial to neutralise the toxin first before attempting to destroy or eliminate the pathogen.
5. Provision of immunity so that you do not get ill if you are infected again with the same pathogen.
The response to infection can be divided into five stages:
1. Awareness of infection. Obviously the body cannot begin to mount a defensive response against a pathogen until it is aware of the presence of the pathogen.
2. The immediate response to infection. This involves the activity of cells and other factors that are present at the time of infection but may require their recruitment to the site of infection and activation once there.
3. The delayed response to infection. If the immediate response to infection is not effective in eliminating the pathogen, the next stage is to generate new cells and factors to deal with the infection.
4. Destruction or elimination of the pathogen or neutralisation of the threat posed by pathogens. The optimal way of dealing with pathogens is to kill them or eliminate them from the body. However, in some cases where pathogens are producing a powerful toxin it may be more beneficial to neutralise the toxin first before attempting to destroy or eliminate the pathogen.
5. Provision of immunity so that you do not get ill if you are infected again with the same pathogen.
вторник, 12 октября 2010 г.
Summary
• The body is continually exposed to infectious organisms that have the potential to cause disease (pathogens). Most pathogens are prevented from entering the body by a combination of physical, chemical and biochemical defence mechanisms. However, some pathogens can breach the barriers and in some cases the barriers are breached by injury or other causes.
• Pathogens vary enormously in terms of size, ways in which they enter the body, how they multiply, whether they replicate intra- or extracellularly, replication rates, mechanisms by which they spread through the body and ways in which they actually cause disease.
• The variety of pathogenic lifestyles means that the immune system must have an equally varied repertoire of mechanisms for dealing with the diversity of threats.
• Pathogens vary enormously in terms of size, ways in which they enter the body, how they multiply, whether they replicate intra- or extracellularly, replication rates, mechanisms by which they spread through the body and ways in which they actually cause disease.
• The variety of pathogenic lifestyles means that the immune system must have an equally varied repertoire of mechanisms for dealing with the diversity of threats.
понедельник, 11 октября 2010 г.
EXAMPLES OF THE LIFESTYLE OF PATHOGENIC ORGANISMS.
Measles.
The measles virus enters the body through the respiratory tract. It then travels to local lymph nodes and lymphoid tissue located in the mucosa. After a few days the virus spreads to other lymphoid tissue, including the spleen, where it begins to replicate. After a week or so, large quantities of the virus spread via the bloodstream to epithelial sites throughout the body. The presence of large amounts of virus at these sites gives rise to the various symptoms seen in measles. Virus in the respiratory tract causes runny nose and coughing. There is inflammation of the conjunctiva, and the presence of the virus in the skin causes the characteristic rash seen in measles.
Typhoid.
If the Salmonella typhi bacterium is ingested and the dose is big enough, some bacteria will survive the acid environment of the stomach and enter the intestine. Bacteria penetrate the gut mucosa through specialised lymphoid structures known as Peyer’s patches and spread to the intestinal lymph nodes, where they proliferate in macrophages. Eventually the organisms reach the bloodstream, where they spread mainly to the liver, bone marrow and spleen, where they continue to multiply. This results in a further large increase in bacterial numbers and subsequent spread of the organism to other tissues such as the kidney and the gall bladder via blood or the biliary tract. The bacteria can also spread to the brain, heart and skin. The bacteria then invade the intestinal tract in much larger numbers than seen with the original infection and cause inflammatory lesions in the Peyer’s patches, which may result in ulceration of the intestinal wall. The presence of the bacteria in other sites may cause meningitis, osteomyelitis, endocarditis and rashes.
The measles virus enters the body through the respiratory tract. It then travels to local lymph nodes and lymphoid tissue located in the mucosa. After a few days the virus spreads to other lymphoid tissue, including the spleen, where it begins to replicate. After a week or so, large quantities of the virus spread via the bloodstream to epithelial sites throughout the body. The presence of large amounts of virus at these sites gives rise to the various symptoms seen in measles. Virus in the respiratory tract causes runny nose and coughing. There is inflammation of the conjunctiva, and the presence of the virus in the skin causes the characteristic rash seen in measles.
Typhoid.
If the Salmonella typhi bacterium is ingested and the dose is big enough, some bacteria will survive the acid environment of the stomach and enter the intestine. Bacteria penetrate the gut mucosa through specialised lymphoid structures known as Peyer’s patches and spread to the intestinal lymph nodes, where they proliferate in macrophages. Eventually the organisms reach the bloodstream, where they spread mainly to the liver, bone marrow and spleen, where they continue to multiply. This results in a further large increase in bacterial numbers and subsequent spread of the organism to other tissues such as the kidney and the gall bladder via blood or the biliary tract. The bacteria can also spread to the brain, heart and skin. The bacteria then invade the intestinal tract in much larger numbers than seen with the original infection and cause inflammatory lesions in the Peyer’s patches, which may result in ulceration of the intestinal wall. The presence of the bacteria in other sites may cause meningitis, osteomyelitis, endocarditis and rashes.
воскресенье, 10 октября 2010 г.
Conclusion
From the above description of the variety of pathogens and the way they live and cause disease, it can be appreciated that the immune system is faced with an enormous variety of problems when trying to protect the body from disease caused by all the different types of pathogens. There is one additional factor that further challenges the immune system and increases the complexity of the immune responses required. Pathogens have coevolved with the immune system and have developed survival strategies to counter attempts to eliminate them. It is obviously in the pathogen’s best interests to survive in the host, and natural selection occurs so that pathogens with an improved ability to survive and multiply within hosts will have a selective advantage and become more common. The evolution of some pathogens seems to have been strongly influenced by the need to evade the immune response; for instance, cytomegalovirus (a cause of pneumonia) has devoted 30% of its genome to subverting the immune response against it. The immune system has accordingly had to evolve an equally complex variety of mechanisms to deal with the wide range of threats posed by different pathogens.
пятница, 8 октября 2010 г.
TOXINS.
Exotoxins.
Exotoxins are secreted products, usually of bacteria but sometimes protozoa and fungi. They can act in a number of ways:
• Inhibition of protein synthesis. Corynebacterium diphtheriae, the cause of diphtheria, produces a toxin that causes ADP-ribosylation of elongation factor-2, thereby stopping protein synthesis. It is extremely potent and one molecule of toxin is capable of killing a cell. Escherichia coli, Vibrio cholerae and Bordatella pertussis (the cause of whooping cough) also produce toxins that cause ADPribosylation of proteins.
Toxins from Shigella dysenteriae and E. coli strain O157:H7 (a cause of dangerous food poisoning) inhibit protein synthesis by removing adenine from 28s rRNA.
• Increase in cAMP. A number of bacteria produce toxins that raise cAMP levels. These include V. cholera, Bacillus anthracis, B. pertussis and some strains of E. coli. The consequence of increased cAMP levels is alterations in ion transport and hence fluid movement, often resulting in severe oedema.
• Neurotoxins. Members of the Clostridium family produce particularly potent neurotoxins. Clostridium tetani produces a toxin that prevents the release of glycine, an inhibitory neurotransmitter. This results in overactivity and muscle spasm including the typical lockjaw. Clostridium botulinum produces a neurotoxin that stimulates release of acetyl choline, leading to paralysis. It is one of the most potent toxins known and it is estimated that less than 1μg can kill a person.
• Enzymes that disrupt cell walls. Clostridium perfringens, a cause of gas gangrene, produces a toxin called α-toxin, which is a phospholipase that hydrolyses lecithin in the cell membrane, resulting in cell death.
• Superantigens. Some bacteria, particularly Staphylococcus and Streptococcus, produce toxins that cause excessive stimulation of the immune system (specifically of T lymphocytes; see Chapter 6). This leads to the production of factors by the immune system that cause the symptoms of shock. Toxic shock syndrome and food poisoning are two consequences of these toxins.
Exotoxins are secreted products, usually of bacteria but sometimes protozoa and fungi. They can act in a number of ways:
• Inhibition of protein synthesis. Corynebacterium diphtheriae, the cause of diphtheria, produces a toxin that causes ADP-ribosylation of elongation factor-2, thereby stopping protein synthesis. It is extremely potent and one molecule of toxin is capable of killing a cell. Escherichia coli, Vibrio cholerae and Bordatella pertussis (the cause of whooping cough) also produce toxins that cause ADPribosylation of proteins.
Toxins from Shigella dysenteriae and E. coli strain O157:H7 (a cause of dangerous food poisoning) inhibit protein synthesis by removing adenine from 28s rRNA.
• Increase in cAMP. A number of bacteria produce toxins that raise cAMP levels. These include V. cholera, Bacillus anthracis, B. pertussis and some strains of E. coli. The consequence of increased cAMP levels is alterations in ion transport and hence fluid movement, often resulting in severe oedema.
• Neurotoxins. Members of the Clostridium family produce particularly potent neurotoxins. Clostridium tetani produces a toxin that prevents the release of glycine, an inhibitory neurotransmitter. This results in overactivity and muscle spasm including the typical lockjaw. Clostridium botulinum produces a neurotoxin that stimulates release of acetyl choline, leading to paralysis. It is one of the most potent toxins known and it is estimated that less than 1μg can kill a person.
• Enzymes that disrupt cell walls. Clostridium perfringens, a cause of gas gangrene, produces a toxin called α-toxin, which is a phospholipase that hydrolyses lecithin in the cell membrane, resulting in cell death.
• Superantigens. Some bacteria, particularly Staphylococcus and Streptococcus, produce toxins that cause excessive stimulation of the immune system (specifically of T lymphocytes; see Chapter 6). This leads to the production of factors by the immune system that cause the symptoms of shock. Toxic shock syndrome and food poisoning are two consequences of these toxins.
вторник, 5 октября 2010 г.
Pathogenesis
The final stage of the disease process (although it may not be the final stage of the infection) is the actual production of disease. Many microorganisms live in or on the body without causing disease. These organisms are called commensal organisms and may be beneficial to the host: the production of lactic and proprionic acids by lactobacilli in the vagina inhibits the growth of many other bacteria and many commensal organisms compete with pathogens for ‘living space’ in the gut. Pathogens differ in that they cause disease by one or more mechanisms. These include the following:
• Secretion of toxins. Many organisms, especially bacteria, secrete toxins that either directly or indirectly account for most of the pathology caused by the organism. These include the powerful neurotoxins secreted by the Clostridium family of bacteria responsible for tetanus or botulism food poisoning, toxins of the bacteria Shigella dysenteriae and Vibrio cholerae that cause dysentery and cholera respectively, and toxins secreted by Streptococcus pyogenes, which can cause scarlet fever. Some protozoa and fungi also secrete exotoxins.
• Endotoxins. Endotoxins, rather than being secreted, are components of the cell wall of pathogens. They are particularly prevalent in Gramnegative bacteria (e.g. Salmonella) but are also found in other bacteria, some yeasts and protozoa. Unlike exotoxins, which have direct, very specific toxic effects, endotoxins act by causing cells of the host to produce factors that cause fever, a fall in blood pressure and other symptoms.

• Direct killing of host cells. Some intracellular dwelling pathogens replicate within cells and leave the cells (usually by budding from the cell surface) with relatively little damage to the cell. This results in the continuous production of infectious particles by an infected cell. Other pathogens replicate within the cell and kill the cell, which bursts open (a process called cell lysis), thereby releasing many infectious particles. Many viruses and protozoa lyse host cells in this way; if this lysis is extensive enough, it will result in disease.
• Physical blockage. Larger pathogens may cause pathology purely by their physical presence. Probably the most dramatic example of this is elephantiasis caused by the filarial worms. By blocking lymphatic drainage these organisms can cause massive swelling of the breasts, testes and legs.
• Secretion of toxins. Many organisms, especially bacteria, secrete toxins that either directly or indirectly account for most of the pathology caused by the organism. These include the powerful neurotoxins secreted by the Clostridium family of bacteria responsible for tetanus or botulism food poisoning, toxins of the bacteria Shigella dysenteriae and Vibrio cholerae that cause dysentery and cholera respectively, and toxins secreted by Streptococcus pyogenes, which can cause scarlet fever. Some protozoa and fungi also secrete exotoxins.
• Endotoxins. Endotoxins, rather than being secreted, are components of the cell wall of pathogens. They are particularly prevalent in Gramnegative bacteria (e.g. Salmonella) but are also found in other bacteria, some yeasts and protozoa. Unlike exotoxins, which have direct, very specific toxic effects, endotoxins act by causing cells of the host to produce factors that cause fever, a fall in blood pressure and other symptoms.

Damage caused by microbial exotoxins and endotoxins. Many pathogens secrete exotoxins, which bind to molecules on host cells, enter the host cell and kill it. Endotoxins are components of the cell wall of some pathogens. Endotoxins in the cell wall, or shed endotoxin, bind to receptors on certain host cells (such as macrophages) and stimulate the host cells to secrete products that damage neighbouring host cells and tissue.
• Direct killing of host cells. Some intracellular dwelling pathogens replicate within cells and leave the cells (usually by budding from the cell surface) with relatively little damage to the cell. This results in the continuous production of infectious particles by an infected cell. Other pathogens replicate within the cell and kill the cell, which bursts open (a process called cell lysis), thereby releasing many infectious particles. Many viruses and protozoa lyse host cells in this way; if this lysis is extensive enough, it will result in disease.
• Physical blockage. Larger pathogens may cause pathology purely by their physical presence. Probably the most dramatic example of this is elephantiasis caused by the filarial worms. By blocking lymphatic drainage these organisms can cause massive swelling of the breasts, testes and legs.
понедельник, 4 октября 2010 г.
Spread of pathogens
The way in which organisms spread through the body is influenced to some extent by whether they live intracellularly, extracellularly or both. Organisms that live extracellularly are able to spread via body fluids such as blood. However, even organisms that replicate intracellularly may be able to leave the cell and spread via an extracellular route. Organisms can spread in the following ways:
• Cell to cell contact. Many organisms, especially viruses, spread directly from cell to cell with essentially no extracellular component to their lifestyle. These pathogens tend to cause localised infections such as seen in influenza, where only the respiratory tract is infected. However, localised infections can still cause widespread symptoms, so that ’flu causes headache, fever and muscle-ache.
• Via blood and lymphatic vessels. The commonest, and fastest, way in which pathogens can spread through the body is via the bloodstream. Since all organs and tissues require a blood supply, microorganisms in the blood have the potential to spread to all sites. However, individual pathogens show a preference to localise in particular organs or tissues that may differ from pathogen to pathogen.
The lymphatic vessels form a circulatory system that parallels that of the blood. There are important differences between the two systems, however. The circulation of the lymphatic fluid is maintained not by the heart but by the movement of the muscles surrounding the lymphatic vessels; thus lymphatic fluid flows at a much more sluggish rate than blood. Moreover, tissue fluid can drain directly into lymphatic vessels. Organisms can easily enter lymphatic vessels draining the site of infection, where they will be conveyed to the local lymph nodes.
• Spread via body cavities. Microorganisms that have infected one organ in a body cavity such as the peritoneum may occasionally spread via the cavity to other organs located within it.
• Spread via nervous system. This is a particularly important route of spread for certain viruses. Viruses can spread via peripheral nerves to the central nervous system (CNS) or vice versa. In some instances this route of spread allows the virus to become more widespread within the nervous system where it resides and causes disease (e.g. herpes simplex virus). In other cases the virus travels via nerves to infect other organs. The rabies virus infects the salivary glands in this way, enabling the virus present in the saliva to be transmitted via a bite.
• Cell to cell contact. Many organisms, especially viruses, spread directly from cell to cell with essentially no extracellular component to their lifestyle. These pathogens tend to cause localised infections such as seen in influenza, where only the respiratory tract is infected. However, localised infections can still cause widespread symptoms, so that ’flu causes headache, fever and muscle-ache.
• Via blood and lymphatic vessels. The commonest, and fastest, way in which pathogens can spread through the body is via the bloodstream. Since all organs and tissues require a blood supply, microorganisms in the blood have the potential to spread to all sites. However, individual pathogens show a preference to localise in particular organs or tissues that may differ from pathogen to pathogen.
The lymphatic vessels form a circulatory system that parallels that of the blood. There are important differences between the two systems, however. The circulation of the lymphatic fluid is maintained not by the heart but by the movement of the muscles surrounding the lymphatic vessels; thus lymphatic fluid flows at a much more sluggish rate than blood. Moreover, tissue fluid can drain directly into lymphatic vessels. Organisms can easily enter lymphatic vessels draining the site of infection, where they will be conveyed to the local lymph nodes.
• Spread via body cavities. Microorganisms that have infected one organ in a body cavity such as the peritoneum may occasionally spread via the cavity to other organs located within it.
• Spread via nervous system. This is a particularly important route of spread for certain viruses. Viruses can spread via peripheral nerves to the central nervous system (CNS) or vice versa. In some instances this route of spread allows the virus to become more widespread within the nervous system where it resides and causes disease (e.g. herpes simplex virus). In other cases the virus travels via nerves to infect other organs. The rabies virus infects the salivary glands in this way, enabling the virus present in the saliva to be transmitted via a bite.
воскресенье, 3 октября 2010 г.
Multiplication of pathogens
Most initial infections are local, i.e. the infectious agent gains entry to the body at a single site, e.g. via an insect bite or infection of a particular mucosal surface. The next stages of infection involve multiplication and spread of the pathogen. These can be considered part of the lifestyle of the pathogen, and infectious organisms vary enormously in lifestyle.
Multiplication of pathogens provides variety at three levels: the mode of multiplication, the site of replication and the rate of multiplication.
Mode of multiplication.
Different pathogens multiply in very different ways. Many single-celled organisms, including bacteria, yeasts and protozoan parasites, divide by simple cell division. Viruses, however, have a completely different mode of multiplication called replication. Following infection of a cell, viral particles disassemble and, under direction of viral nucleic acid (DNA or RNA), new viral proteins and genetic material are synthesised. Eventually new viral particles are assembled and leave the cell. This can occur by the cell bursting open and releasing viral particles to infect other cells, resulting in cell lysis and death of the cell. Alternatively the cell can shed viral particles in a more gradual manner, a process known as budding, which does not result in the death of the cell. Finally many parasitic worms do not multiply directly but lay eggs, which provide additional sources of infection for other organisms.
Site of replication.
Pathogens can live and multiply inside host cells or outside the cells. Many bacteria, yeasts and parasites multiply extracellularly. Viruses by their nature have to replicate intracellularly because they lack enzymes and other cofactors necessary for synthesising viral proteins. Many bacteria and protozoan parasites also replicate intracellularly. Some organisms can live in either an intracellular or an extracellular environment (e.g. Mycobacterium tuberculosis, Neisseria gonorrhoeae). Parasites (e.g. trypanosomes) have the most complicated life cycles, which can often involve both an intracellular and extracellular stage.
The site in which pathogens live and multiply poses different problems for the immune system. The most important of these is whether the pathogen has an intracellular stage, because during this stage the organism may be partially hidden from the immune system. However, as we shall, the immune system has even evolved ways of detecting whether infected host cells are harbouring hidden pathogens.

Rates of multiplication.
The time taken for pathogens to reproduce themselves varies enormously. Some bacteria under optimal conditions in vitro can divide every 20 minutes. At this rate of division a single bacterium would produce over 1021 progeny in a day! Obviously this rate of replication is unsustainable for long, even under optimal in vitro conditions, and it is debatable whether it is ever reached in vivo. Viral replication can result in hundreds or thousands of progeny being produced from a single virion in hours. Other pathogens have low rates of replication. Not all bacteria have the capacity to divide rapidly and some, such as the mycobacteria, the causes of tuberculosis and leprosy, have a doubling time of many days. Some parasitic worms never replicate within the host, although they may lay eggs, thereby increasing the number of organisms that can infect other hosts. However, again the rate of egg laying can vary enormously: Schistosoma mansoni, the cause of the disease schistosomiasis, lays only 200 eggs a day while Ascaris lumbricoides, a roundworm, may lay over 200 000.
Multiplication of pathogens provides variety at three levels: the mode of multiplication, the site of replication and the rate of multiplication.
Mode of multiplication.
Different pathogens multiply in very different ways. Many single-celled organisms, including bacteria, yeasts and protozoan parasites, divide by simple cell division. Viruses, however, have a completely different mode of multiplication called replication. Following infection of a cell, viral particles disassemble and, under direction of viral nucleic acid (DNA or RNA), new viral proteins and genetic material are synthesised. Eventually new viral particles are assembled and leave the cell. This can occur by the cell bursting open and releasing viral particles to infect other cells, resulting in cell lysis and death of the cell. Alternatively the cell can shed viral particles in a more gradual manner, a process known as budding, which does not result in the death of the cell. Finally many parasitic worms do not multiply directly but lay eggs, which provide additional sources of infection for other organisms.
Site of replication.
Pathogens can live and multiply inside host cells or outside the cells. Many bacteria, yeasts and parasites multiply extracellularly. Viruses by their nature have to replicate intracellularly because they lack enzymes and other cofactors necessary for synthesising viral proteins. Many bacteria and protozoan parasites also replicate intracellularly. Some organisms can live in either an intracellular or an extracellular environment (e.g. Mycobacterium tuberculosis, Neisseria gonorrhoeae). Parasites (e.g. trypanosomes) have the most complicated life cycles, which can often involve both an intracellular and extracellular stage.
The site in which pathogens live and multiply poses different problems for the immune system. The most important of these is whether the pathogen has an intracellular stage, because during this stage the organism may be partially hidden from the immune system. However, as we shall, the immune system has even evolved ways of detecting whether infected host cells are harbouring hidden pathogens.

Multiplication of pathogens. Many bacteria divide by simple cell fission. Viruses must infect host cells to replicate. Parasitic worms (helminths) often lay eggs, which are transmitted to new hosts before developing into worms.
Rates of multiplication.
The time taken for pathogens to reproduce themselves varies enormously. Some bacteria under optimal conditions in vitro can divide every 20 minutes. At this rate of division a single bacterium would produce over 1021 progeny in a day! Obviously this rate of replication is unsustainable for long, even under optimal in vitro conditions, and it is debatable whether it is ever reached in vivo. Viral replication can result in hundreds or thousands of progeny being produced from a single virion in hours. Other pathogens have low rates of replication. Not all bacteria have the capacity to divide rapidly and some, such as the mycobacteria, the causes of tuberculosis and leprosy, have a doubling time of many days. Some parasitic worms never replicate within the host, although they may lay eggs, thereby increasing the number of organisms that can infect other hosts. However, again the rate of egg laying can vary enormously: Schistosoma mansoni, the cause of the disease schistosomiasis, lays only 200 eggs a day while Ascaris lumbricoides, a roundworm, may lay over 200 000.
суббота, 2 октября 2010 г.
Invasion – entry of pathogens into the body
Routes by which infectious organisms gain entry into the body include the skin, respiratory tract, gastro-intestinal (GI) tract and GU tract. There are fundamentally two ways in which infectious agents cross the physical and chemical barriers: either they are able to penetrate the intact barriers at one or more anatomical sites, or the physical barriers are damaged and breached, allowing entry of the organism.
Penetration of intact skin or mucosa
• Skin. Few organisms are able to penetrate intact skin. However, some parasites (e.g. hookworm) or their larvae (e.g. schistosoma) can do this. Other agents, such as wart viruses, set up infection in the skin and do not enter further into the body.


• Mucosa. Mucosa, being softer and damper than skin, are much more frequent sites of entry and all intact mucosa can be penetrated by some organisms. Examples are shown in table Pathogens can cross epithelia by passing through epithelial cells, as in the case of the meningococcus (a bacterium causing meningitis), or by passing between the epithelial cells, seen with Haemophilus influenzae.
Penetration of damaged skin or mucosa.
There are many ways in which skin or mucosa can be damaged, allowing entry of infectious organisms that could not cross intact skin or mucosa. Damage to skin is a particularly important route of infection and can occur in a number of ways:
• Burns. Burns, especially severe ones, pose a major risk for infection, particularly with Staphylococcus, Streptococcus, Pseudomonas and Clostridium tetanus.
• Cuts and wounds. These can allow entry of similar organisms to those seen after burns.
• Insect bites. Numerous infections are transmitted via insect bites. These include malaria, typhus and plague.
• Animal bites. Animal bites can provide direct transmission of infection, such as in rabies. Because they cause significant damage to the skin, bites can allow the entry of the same environmental pathogens as burns, cuts and wounds (see above).
• Human behaviour. Various aspects of uniquely human behaviour can result in the skin being penetrated. Sharing of syringes by intravenous (IV) drug users exposes them to risk of hepatitis and human immunodeficiency virus (HIV). A number of viral infections (hepatitis, HIV) have been transmitted by blood transfusion and blood products (e.g. factor VIII for haemophiliacs) before appropriate screening procedures were developed. Transplantation has also resulted in transmission of infection before the introduction of appropriate donor screening.
Damage to mucosa may not increase the likelihood of infection to the same extent as damage to the skin. However, physical or chemical damage may allow entry of some organisms (e.g. smoking increases the risk of respiratory bacterial infections). Furthermore, infection of the mucosa with a virus may cause damage and facilitate the entry of bacterial pathogens.
Penetration of intact skin or mucosa
• Skin. Few organisms are able to penetrate intact skin. However, some parasites (e.g. hookworm) or their larvae (e.g. schistosoma) can do this. Other agents, such as wart viruses, set up infection in the skin and do not enter further into the body.

Entry of pathogens into the body. Insect bites, cuts, burns and animal bites breach the skin barrier, allowing entry of pathogens. Some parasites can penetrate intact skin while many pathogens penetrate intact mucosa of the respiratory, intestinal and genito-urinary tracts.

Mucosal sites of entry for pathogens
• Mucosa. Mucosa, being softer and damper than skin, are much more frequent sites of entry and all intact mucosa can be penetrated by some organisms. Examples are shown in table Pathogens can cross epithelia by passing through epithelial cells, as in the case of the meningococcus (a bacterium causing meningitis), or by passing between the epithelial cells, seen with Haemophilus influenzae.
Penetration of damaged skin or mucosa.
There are many ways in which skin or mucosa can be damaged, allowing entry of infectious organisms that could not cross intact skin or mucosa. Damage to skin is a particularly important route of infection and can occur in a number of ways:
• Burns. Burns, especially severe ones, pose a major risk for infection, particularly with Staphylococcus, Streptococcus, Pseudomonas and Clostridium tetanus.
• Cuts and wounds. These can allow entry of similar organisms to those seen after burns.
• Insect bites. Numerous infections are transmitted via insect bites. These include malaria, typhus and plague.
• Animal bites. Animal bites can provide direct transmission of infection, such as in rabies. Because they cause significant damage to the skin, bites can allow the entry of the same environmental pathogens as burns, cuts and wounds (see above).
• Human behaviour. Various aspects of uniquely human behaviour can result in the skin being penetrated. Sharing of syringes by intravenous (IV) drug users exposes them to risk of hepatitis and human immunodeficiency virus (HIV). A number of viral infections (hepatitis, HIV) have been transmitted by blood transfusion and blood products (e.g. factor VIII for haemophiliacs) before appropriate screening procedures were developed. Transplantation has also resulted in transmission of infection before the introduction of appropriate donor screening.
Damage to mucosa may not increase the likelihood of infection to the same extent as damage to the skin. However, physical or chemical damage may allow entry of some organisms (e.g. smoking increases the risk of respiratory bacterial infections). Furthermore, infection of the mucosa with a virus may cause damage and facilitate the entry of bacterial pathogens.
пятница, 1 октября 2010 г.
Barriers to infection
The body has many physical, chemical and biochemical barriers that make it much more difficult for pathogens to gain entry into the body.

The physical barriers to infection are as follows:
• Skin and mucosa. Intact skin and mucosa provide a physical barrier to prevent entry of organisms.
• Cilia. The respiratory tract is lined with little hair-like structures that beat in such a way as to propel particles towards the throat, where they can be expelled by coughing or swallowing and excretion.
• Mucus. Mucus is secreted by epithelial cells of the gut, respiratory tract and genito-urinary (GU) tract. It has the unusual properties of being sticky and slimy at the same time and is able to entrap microorganisms so they can be expelled. In the respiratory tract, cilia and mucus combine to provide an effective way of trapping and eliminating microbes.
The chemical and biochemical defences are as follows:
• Acids. Hydrochloric acid secreted by the stomach is lethal to many (though not all) bacteria. Commensal bacteria in the vagina produce lactic and proprionic acid resulting in a low pH, which is inhibitory to the division of many bacteria.
• Fatty acids. Sebaceous glands in the skin produce fatty acids that have antimicrobial properties.
• Lysozyme. This is present in sweat, tears and many other secretions. It breaks down peptidoglycans in bacterial cell walls, thus damaging and killing the bacteria.
• Defensins. Defensins are antimicrobial peptides that are found in the secretions of mucosa and skin.
• Cathelicidins. These antibacterial peptides were originally discovered as insect defence peptides. Other members of the cathelicidin family have been found in mucosal secretions.
• Collectins. Collectins are proteins that can bind sugars on microbial surfaces and promote the elimination of microbes. Proteins that bind sugars are known as lectins; because collectins bind sugars in a calciumdependent manner, they are known as C-type lectins. The A and D lung surfactants are collectins that provide protection at the lung surface; other collectins, such as mannose-binding protein, are found in serum.
The physical and chemical barriers are very effective at preventing pathogens from entering the body and they exclude more than 99.9% of the infectious organisms we are exposed to. However, organisms do infect the body. This can occur in a number of ways.

Physical and chemical defence mechanisms.
The physical barriers to infection are as follows:
• Skin and mucosa. Intact skin and mucosa provide a physical barrier to prevent entry of organisms.
• Cilia. The respiratory tract is lined with little hair-like structures that beat in such a way as to propel particles towards the throat, where they can be expelled by coughing or swallowing and excretion.
• Mucus. Mucus is secreted by epithelial cells of the gut, respiratory tract and genito-urinary (GU) tract. It has the unusual properties of being sticky and slimy at the same time and is able to entrap microorganisms so they can be expelled. In the respiratory tract, cilia and mucus combine to provide an effective way of trapping and eliminating microbes.
The chemical and biochemical defences are as follows:
• Acids. Hydrochloric acid secreted by the stomach is lethal to many (though not all) bacteria. Commensal bacteria in the vagina produce lactic and proprionic acid resulting in a low pH, which is inhibitory to the division of many bacteria.
• Fatty acids. Sebaceous glands in the skin produce fatty acids that have antimicrobial properties.
• Lysozyme. This is present in sweat, tears and many other secretions. It breaks down peptidoglycans in bacterial cell walls, thus damaging and killing the bacteria.
• Defensins. Defensins are antimicrobial peptides that are found in the secretions of mucosa and skin.
• Cathelicidins. These antibacterial peptides were originally discovered as insect defence peptides. Other members of the cathelicidin family have been found in mucosal secretions.
• Collectins. Collectins are proteins that can bind sugars on microbial surfaces and promote the elimination of microbes. Proteins that bind sugars are known as lectins; because collectins bind sugars in a calciumdependent manner, they are known as C-type lectins. The A and D lung surfactants are collectins that provide protection at the lung surface; other collectins, such as mannose-binding protein, are found in serum.
The physical and chemical barriers are very effective at preventing pathogens from entering the body and they exclude more than 99.9% of the infectious organisms we are exposed to. However, organisms do infect the body. This can occur in a number of ways.
четверг, 30 сентября 2010 г.
Disease production by pathogens
The first stage of disease production by pathogens is infection, or entry of the pathogen into the body. Nearly all pathogens must gain entry into the body before they can begin to replicate or spread. A few pathogens can exist on the skin (e.g. viruses causing warts) or in the gut (e.g. bacteria causing cholera) without technically entering the body. However, infection is not made easy for pathogens because the body has many physical and chemical barriers to try to prevent pathogens entering the body.
среда, 29 сентября 2010 г.
Stages of disease production by pathogens
Size is not the only way in which infectious organisms vary. They also vary enormously with respect to how they enter and live within the body and actually cause disease. Infection and disease production by pathogenic organisms can be divided into four stages:
1. Invasion.
2. Multiplication.
3. Spread.
4. Production of disease (pathogenesis).
Although infection usually involves all of these steps, there are many exceptions in terms of both the steps involved and their order. Some pathogens do not spread significantly or even technically gain entry to the body. Organisms may replicate locally before spreading or may spread through the body before beginning significant replication. Pathogens show considerable variation at each of these stages of infection, as will be described below.
1. Invasion.
2. Multiplication.
3. Spread.
4. Production of disease (pathogenesis).
Although infection usually involves all of these steps, there are many exceptions in terms of both the steps involved and their order. Some pathogens do not spread significantly or even technically gain entry to the body. Organisms may replicate locally before spreading or may spread through the body before beginning significant replication. Pathogens show considerable variation at each of these stages of infection, as will be described below.
вторник, 28 сентября 2010 г.
Size of pathogens
One feature of the range of pathogenic organisms listed in table is the enormous variation in size. Viruses are the smallest infectious organisms, being 20–400 nm in size. At the other end of the scale some parasitic worms, such as the tapeworm, can be up to 7 m (20 ft) in length. This represents a difference in scale of a factor of 109. To put that into some sort of perspective, if a virus were the size of a tennis ball, a fully developed tapeworm would reach from London to Los Angeles. It does not stretch the imagination too far to appreciate that the problems posed to the immune system by these two organisms would require very different solutions.
понедельник, 27 сентября 2010 г.
Types of pathogen and how they differ
The types of pathogen that can cause disease include many groups of single-celled microorganisms and larger multicellular parasites. Viruses, bacteria, some yeasts, and protozoan parasites are examples of singlecelled pathogens. Fungi and helminths (parasitic worms) are the major multi-cellular pathogens. These pathogens come from very different parts of the biological kingdom and vary considerably in many aspects. Pathogens differ enormously in their size. They also have very different lifestyles and cause disease in a variety of ways.


Size and lifestyle of pathogens
воскресенье, 26 сентября 2010 г.
Why is immunology so complicated?
There are thousands of components to the immune system, and during the course of learning about some of these it can appear that the immune system is far more complicated than necessary for achieving what is, on the surface, the simple task of eliminating an infectious organism. There are a number of reasons why the immune system is complex. The first of these is the desirability of eliminating pathogens without causing damage to the host. Getting rid of a pathogen is theoretically easy. If you had an infection in your liver you could produce a nasty toxin that would kill the pathogen; unfortunately it would also destroy your liver. Killing pathogens is not difficult, but getting rid of pathogens without damaging the host is much more complicated. Imagine if a city in your country was infiltrated by soldiers from another country; you could get rid of the foreign invaders by dropping a nuclear bomb on the city but this would also kill a lot of your compatriots. To go into the city to eliminate or capture the foreign soldiers without causing harm to your compatriots is much more difficult. This analogy raises a major issue concerning the immune system – that of recognition. To eliminate foreign soldiers without killing your own requires that you can tell the two apart. In the same way the immune system must be able to distinguish between pathogens and host cells so that it can direct its destructive powers towards the pathogens. Many of the specialised features of the immune system are involved in recognition of foreign pathogens.
An additional problem facing the immune system is that pathogens come in all shapes and sizes, with different lifestyles and different ways of causing disease. To understand fully the complexity that the immune system must deal with, it is necessary to have some understanding of infectious organisms and the ways in which they cause disease. The rest of this chapter describes how pathogens differ, so that it is possible to get an appreciation of the problems faced by the immune system.
An additional problem facing the immune system is that pathogens come in all shapes and sizes, with different lifestyles and different ways of causing disease. To understand fully the complexity that the immune system must deal with, it is necessary to have some understanding of infectious organisms and the ways in which they cause disease. The rest of this chapter describes how pathogens differ, so that it is possible to get an appreciation of the problems faced by the immune system.
суббота, 25 сентября 2010 г.
The role and complexity of the immune system
The immune system consists of proteins, cells and organs that are concerned with defence of the individual, primarily against the threat of disease caused by infectious organisms. An infectious organism that causes disease is called a pathogen and the individual (person or animal) that is infected by a pathogen is called the host. Not all infectious organisms cause disease and some are actually beneficial, for example bacteria living in the gut help to digest certain foods. Infectious organisms that help the host are called commensal organisms. However, many viruses, bacteria, fungi, yeasts and parasites are pathogenic and we are constantly in danger of infection and disease caused by them.
The importance of the immune system in resistance to infection is demonstrated by the fact that, without treatment, babies born with severe defects of certain components of the immune system die in infancy of overwhelming infection, usually with organisms (e.g. Haemophilus influenzae, Candida albicans or cytomegalovirus) that cause little or no disease in individuals with a normal immune system. More recently, the AIDS (acquired immunodeficiency syndrome) epidemic has brought the consequences of having a poorly functioning immune system to the attention of the public at large. AIDS patients usually die from infection, such as with the yeasts Pneumocystis carinii, which causes pneumonia, or Cryptococcus neo-formans, a cause of meningitis. The immune system normally controls these infections with little or no damage to the host.
The immune system is primarily involved in defence against infectious organisms, but it also plays a role in various other diseases. Evidence is accumulating that the immune system can provide protection against some tumours. Exciting new developments also suggest that immune responses can be induced against tumours that normally do not provoke an immune response. These so-called tumour vaccines offer hope of additional weapons in the armoury against cancer. The immune system is also responsible for the rejection of transplants. In this instance the immune system is acting normally in trying to defend the body against a foreign invader, even though the ‘invader’ is beneficial. Rejection of transplants is a major cause of graft loss and much effort is being devoted to try to prevent transplant rejection.
Although a properly functioning immune system is essential for good health, like any other physiological system the immune system can malfunction and actually cause diseases. The most common form of disease caused by the immune system is allergy, for example hayfever and food allergy, which is caused by an inappropriate immune response against generally harmless material such as pollen or food. Another category of diseases caused by malfunction of the immune system are the autoimmune diseases, including rheumatoid arthritis and some types of diabetes. Autoimmune diseases occur when the immune system attacks the body’s own tissue. Although these diseases caused by the immune system can be threatening and sometimes fatal, on balance a properly functioning immune system is essential for life in the world we inhabit, which is full of disease-causing organisms, some of which, as mentioned above, are invariably fatal if the system does not operate properly.

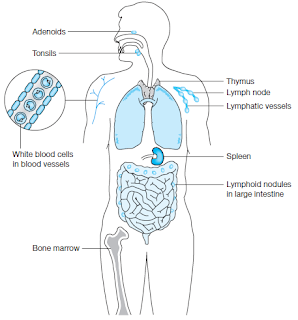
Some components of the immune system. The bone marrow and thymus are
sites of production of lymphocytes, white blood cells involved in immune responses.
Adenoids, tonsils, lymph nodes, spleen and lymphoid nodules are sites where immune
responses are generated. Lymphatic vessels are similar to blood vessels and allow cells of the
immune system to travel throughout the body.
sites of production of lymphocytes, white blood cells involved in immune responses.
Adenoids, tonsils, lymph nodes, spleen and lymphoid nodules are sites where immune
responses are generated. Lymphatic vessels are similar to blood vessels and allow cells of the
immune system to travel throughout the body.
The importance of the immune system in resistance to infection is demonstrated by the fact that, without treatment, babies born with severe defects of certain components of the immune system die in infancy of overwhelming infection, usually with organisms (e.g. Haemophilus influenzae, Candida albicans or cytomegalovirus) that cause little or no disease in individuals with a normal immune system. More recently, the AIDS (acquired immunodeficiency syndrome) epidemic has brought the consequences of having a poorly functioning immune system to the attention of the public at large. AIDS patients usually die from infection, such as with the yeasts Pneumocystis carinii, which causes pneumonia, or Cryptococcus neo-formans, a cause of meningitis. The immune system normally controls these infections with little or no damage to the host.
The immune system is primarily involved in defence against infectious organisms, but it also plays a role in various other diseases. Evidence is accumulating that the immune system can provide protection against some tumours. Exciting new developments also suggest that immune responses can be induced against tumours that normally do not provoke an immune response. These so-called tumour vaccines offer hope of additional weapons in the armoury against cancer. The immune system is also responsible for the rejection of transplants. In this instance the immune system is acting normally in trying to defend the body against a foreign invader, even though the ‘invader’ is beneficial. Rejection of transplants is a major cause of graft loss and much effort is being devoted to try to prevent transplant rejection.
Although a properly functioning immune system is essential for good health, like any other physiological system the immune system can malfunction and actually cause diseases. The most common form of disease caused by the immune system is allergy, for example hayfever and food allergy, which is caused by an inappropriate immune response against generally harmless material such as pollen or food. Another category of diseases caused by malfunction of the immune system are the autoimmune diseases, including rheumatoid arthritis and some types of diabetes. Autoimmune diseases occur when the immune system attacks the body’s own tissue. Although these diseases caused by the immune system can be threatening and sometimes fatal, on balance a properly functioning immune system is essential for life in the world we inhabit, which is full of disease-causing organisms, some of which, as mentioned above, are invariably fatal if the system does not operate properly.
Подписаться на:
Сообщения (Atom)